
はじめに
下部尿路(lower urinary tract;LUT)の機能といえば、私たちは単純に「尿を溜め、出す」という水道設備のような物理的プロセスを想起しがちです。しかし、近年の神経科学と分子生物学の進展は、この一見単純なプロセスが、驚くほど緻密で動的なシグナルネットワークによって支配されていることを明らかにしました。本稿では、最新のレビュー論文(ナラティブレビュー)に基づき、尿路上皮という知られざるセンサー、神経回路の階層的制御、そして尿道という第2の主役が織りなす統合的な生理機能を解き明かします。
尿路上皮:単なる障壁を超えた活動的センサー
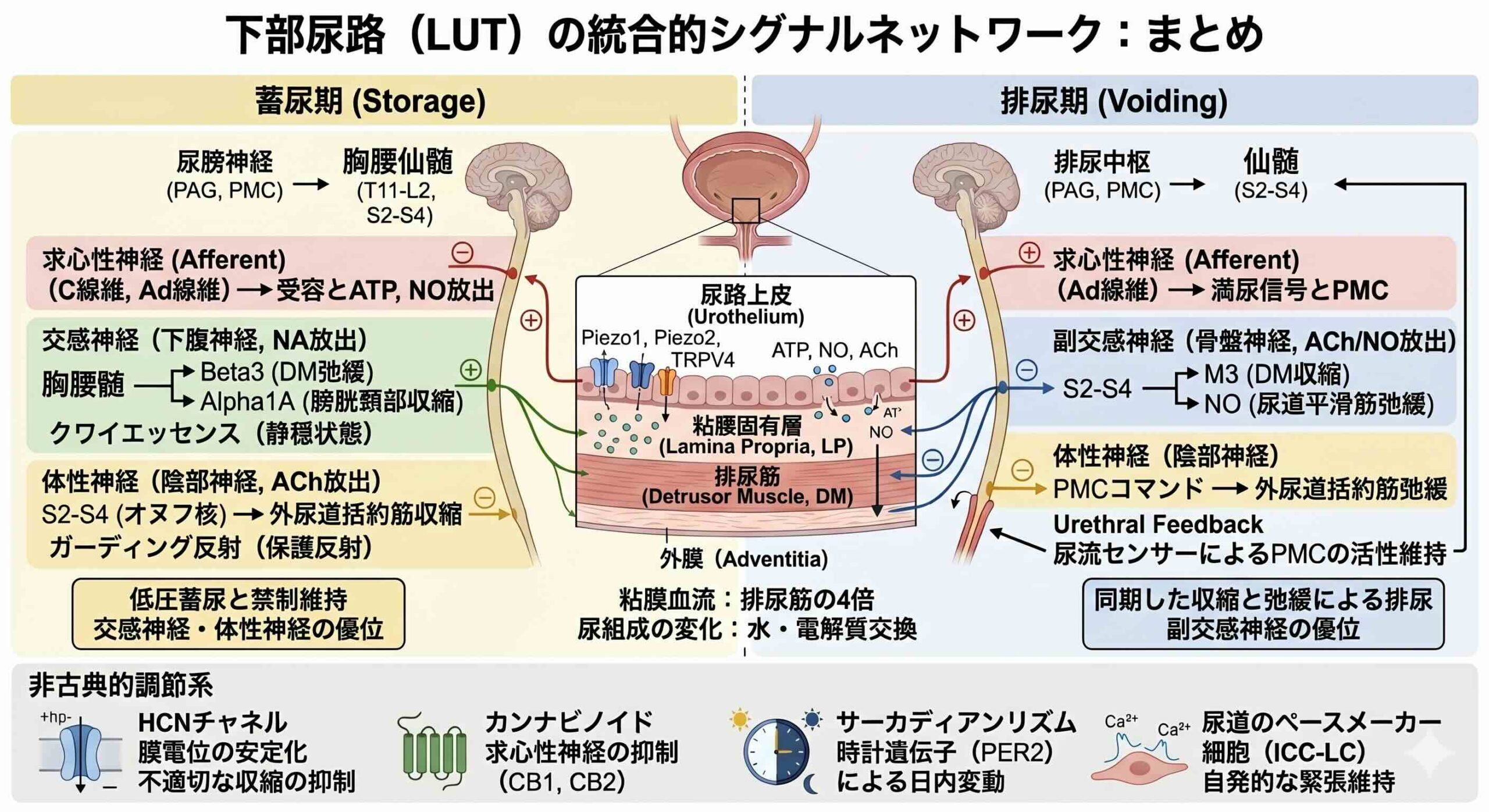
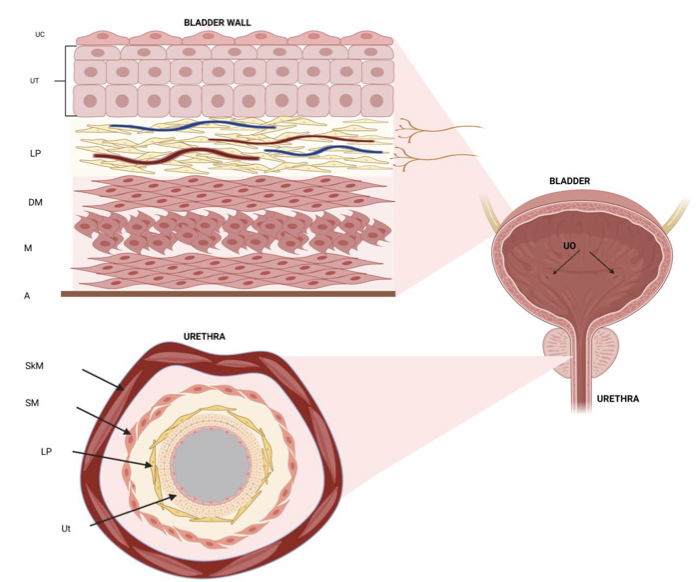
膀胱壁は、内側から尿路上皮、粘膜固有層、排尿筋、外膜の4層で構成されています。かつて尿路上皮は、尿が組織に浸透するのを防ぐだけの受動的な障壁と考えられてきました。しかし、現代の生理学において、尿路上皮は下部尿路の最も重要な感覚インターフェースとしての地位を確立しています。基底細胞、中間細胞、そして最表層のアンブレラ細胞(umbrella cells;UC)という3層構造から成るこの組織は、物理的な伸展や化学的刺激を電気的・化学的な信号へと変換する驚異的なトランスデューサー(変換器)です。
特に注目すべきは、アンブレラ細胞に発現するメカノセンシティブ(機械刺激受容)チャネルの存在です。Piezo1やPiezo2、そしてTRPV4といったチャネルは、膀胱壁のわずかな張力を感知し、細胞内カルシウム濃度を上昇させます。これが引き金となり、アデノシン三リン酸(ATP)、一酸化窒素(NO)、アセチルコリン(ACh)といった神経伝達物質が放出されます。
興味深いことに、膀胱粘膜(尿路上皮と粘膜固有層)は、その高い代謝活性を支えるために、排尿筋よりも約4倍も高い血流を受けています。この潤沢な灌流が、メディエーターの持続的な合成と放出を可能にし、求心性神経の感度を精密に維持しているのです。
分子レベルで展開される情報の増幅と抑制
尿路上皮から放出されたATPは、求心性C線維やAデルタ線維に発現するP2X3受容体に結合し、急速な膜の脱分極を引き起こします。これが脳へと伝わる「尿意」の第一歩となります。また、P2Y受容体を介したオートクリン・パラクリン的なフィードバックループにより、カルシウム信号が増幅され、さらなるATP放出を誘発する仕組みも備わっています。
一方で、過剰な興奮を抑えるブレーキ役も存在します。一酸化窒素(NO)は、求心性神経の興奮性を抑制し、蓄尿期に不適切な排尿反射が起こらないようクワイエッセンス quiescence(静穏状態)を保ちます。
また、近年注目されているのがHCNチャネル(Hyperpolarization-activated cyclic nucleotide–gated (HCN) Channels 過分極活性化環状ヌクレオチド依存性チャネル)です。HCN1からHCN4までのアイソフォームが尿路上皮や神経末端に分布しており、静止膜電位を安定させることで、膀胱の過剰な自発収縮を抑制する役割を果たしています。このHCNチャネルの発現が加齢とともに低下することが、高齢者における過活動膀胱の一因である可能性も示唆されています。
神経回路の階層:脊髄反射から高次脳中枢まで
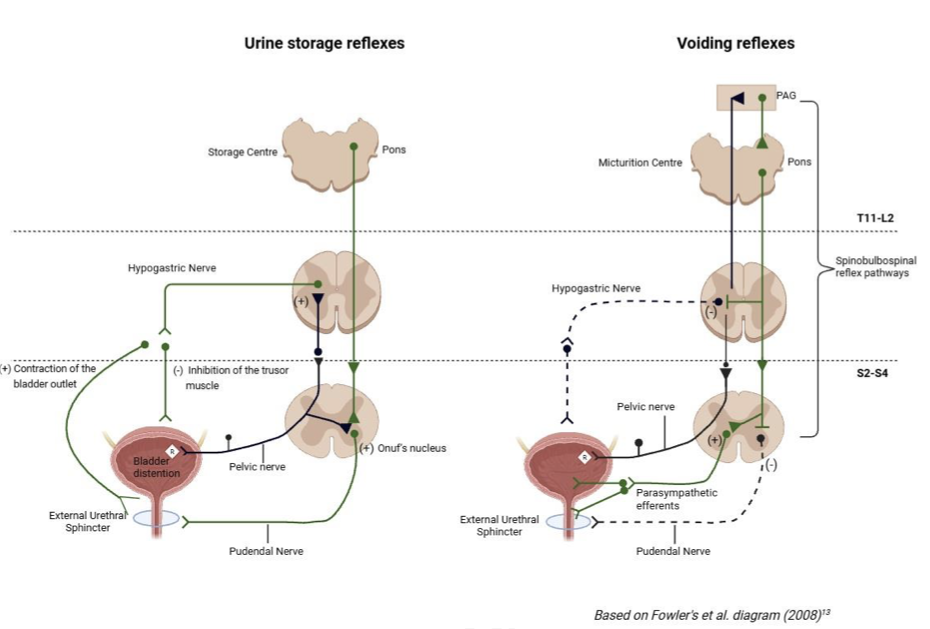
下部尿路の制御は、脊髄レベルの局所反射と、脳幹および大脳皮質による高次制御の二段階で構成されています。
蓄尿期には、膀胱からの微弱な信号が脊髄S2からS4レベルに到達し、ガーディング反射guarding reflex(保護反射)を誘発します。これにより、オヌフ核から発せられる体性神経(陰部神経)の活動が高まり、外尿道括約筋が持続的に収縮します。同時に、胸腰髄レベル(T11からL2)からの交感神経(下腹神経)が、β3受容体を介して排尿筋を弛緩させ、α1A受容体を介して膀胱頸部を収縮させることで、低圧での蓄尿を実現します。
尿が限界まで溜まると、信号は中脳の水道周囲灰白質(periaqueductal gray ;PAG)へと送られます。PAGは、感情や社会的状況、環境情報を統合し、今排尿すべきかどうかを判断するゲートキーパーとして機能します。適切なタイミングと判断されると、橋排尿中枢( pontine micturition centre ;PMC)が活性化し、一転して副交感神経(骨盤神経)の活動が優位になります。ムスカリンM3受容体を介した排尿筋の力強い収縮と、尿道括約筋の弛緩が完璧に同期し、スムーズな排尿が行われるのです。驚くべきことに、これらの感覚入力の約20%は脊髄で反対側に交叉し、両側性の強調的な反射を可能にしています。
尿道:continence(禁制)を支えるアクティブな主役
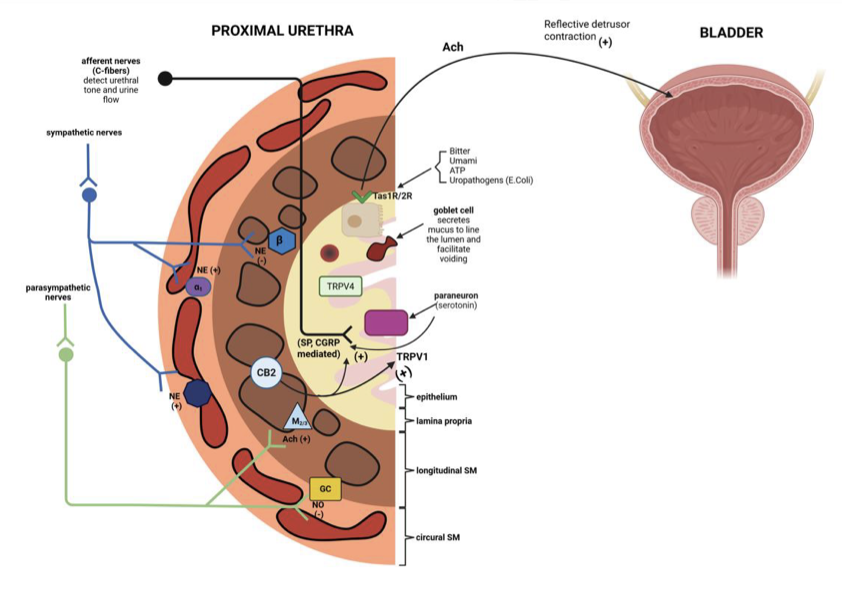
排尿機能の議論では膀胱が主役になりがちですが、本論文は尿道の能動的な役割についても深く掘り下げています。尿道は単なる管ではなく、高度に発達した感覚・運動系を備えた精密なバルブです。
尿道平滑筋には、カハール介在細胞様細胞( Interstitial cells of Cajal-like cells;ICC-LC)と呼ばれる特殊な細胞が存在します。これらの細胞は、リアノジン受容体やIP3受容体を介した自発的なカルシウム波を発生させるペースメーカー機能を持ち、神経入力がなくても尿道の基礎的な緊張状態(トーヌス)を維持することに寄与しています。
性差についても重要な知見があります。女性の尿道は約4センチメートルと短く、その全長の約60%を横紋筋である外尿道括約筋が占めています。一方、男性の尿道は4つのセグメントに分かれ、膀胱頸部には発達した内尿道括約筋が存在し、射精時の精液の逆流を防ぐ役割も兼ねています。このように、構造的な違いがシグナル伝達のパターンにも反映されており、治療戦略を立てる上での重要な考慮事項となります。
時間軸と化学軸の統合:サーカディアンリズムと尿組成の動的変化
下部尿路の機能は、24時間の時間軸の中でもダイナミックに変化します。PER2などの時計遺伝子が尿路上皮や排尿筋に発現しており、明暗サイクルの12時間目付近で発現のピークを迎えます。これにより、夜間の尿産生量や膀胱容量が調節されています。
さらに、驚くべき事実は、尿は膀胱に溜まっている間もその組成が変化し続けているということです。ヒトの尿路上皮は完全な不浸透性ではなく、特に低流量や長時間の貯留時には、水や電解質の交換が行われています。尿中のクレアチニン濃度が単なる希釈の指標として信頼できない場合があるのは、この尿路上皮を介した再吸収や拡散が起こっているためです。膀胱は単なる貯留槽ではなく、尿という環境に対して能動的に応答し、その情報を自身のネットワークに還元しているのです。
研究の新規性と限界(Limitation)
本研究の最大の新規性は、膀胱中心的な視点を脱却し、尿道、神経系、そして非古典的な伝達物質(HCNチャネルやカンナビノイドなど)を網羅的に統合したネットワーク・フレームワークを提示した点にあります。個別の経路をカタログ化するのではなく、各組織がどのように「クロストーク(相互作用)」しているかを整理した点は、今後のベンライ下部尿路疾患(LUTS)の研究に新たな座標軸を与えます。
しかし、いくつかの限界も存在します。解説されているメカニズムの多くは動物モデルや生体外(ex vivo)の標本から得られた知見であり、受容体の発現パターンやICC-LCの寄与度など、種差による不確実性が残っています。また、それぞれのシグナル系が時間軸の中でどのように相互作用し、急性的な反応が長期的な遺伝子発現の変化へと繋がっていくのかについては、未解明な部分が多く、さらなる検証が求められます。
実践への還元:明日から活かせる知見
この統合的シグナルネットワークの理解は、日常の健康管理や臨床的視点にどう活かせるでしょうか。
1つ目は、蓄尿状態の重要性です。生理的な速度での膀胱充満は、急速な充満よりも感覚神経を適切に感作させることが示唆されています。急激な水分摂取は膀胱壁の急峻な伸展を招き、シグナルネットワークを過剰に刺激して尿意切迫感を引き起こしやすくします。穏やかな水分補給が、健全なシグナルバランスを保つ鍵となります。
2つ目は、体内時計との調和です。時計遺伝子が膀胱の感度を支配しているため、不規則な睡眠や生活リズムは、この「尿路の時計」を狂わせ、夜間頻尿や蓄尿障害を悪化させる物理的な要因となり得ます。
3つ目は、感覚の「再学習」です。尿意は、尿路上皮、平滑筋、そして脳の複雑な対話の結果です。骨盤底筋トレーニングなどを通じて、ガーディング反射を能動的に意識することは、中枢神経系におけるゲート制御を強化し、シグナルネットワークの過敏性を鎮めるための生理学的な理にかなった行動と言えます。
私たちの下部尿路は、今日も静かに、しかし熱心に、体内の情報を読み取り、最適な排尿のタイミングを計っています。その複雑な囁きに耳を傾けることは、自身の健康と向き合う新たな視点を与えてくれるはずです。
参考文献
Loufopoulos, I., Papaeftstathiou, E., Kontos, A., & Apostolidis, A. (2026). Integrated signalling networks in the healthy lower urinary tract: A narrative review. Autonomic Neuroscience: Basic and Clinical, 103420. https://doi.org/10.1016/j.autneu.2026.103420

